搬運自微信公衆號攀緣的井蛙
上個世紀80年代,我正是一個孩子。父母有時候會把我帶到他們的“單位”一起喫食堂,省得回家再做飯了。喫飯之外的時間,我就要識相地找個地方待著,別干擾大人工作。“單位”裏能放心讓學前幼兒自由探索的地方,莫過於圖書室了。我經常一個人在幾排書架間爬上爬下,尋找圖多字少的書本報紙,一翻就是大半天。
你別說,還真就給我翻到了寶貝。那是一本薄薄的畫冊,每一頁都畫着一些古生物彩圖:有水母,有三葉蟲,有恐龍,還有猛獁象。圖片全是淡雅的水彩畫風格,生物形象也是早期的復原結構,比如雙足恐龍都是直立身體尾巴拖在地上。在那個匱乏的時代,這本畫冊讓我愛不釋手,每次去“單位”都直奔圖書室,如飢似渴地看,後來更是拿紙筆一張一張地照着描畫。
差不多就是這樣的風格,不過是彩色的,而且大得多。照片攝自[1]。
直到今天,那些古生物的樣子還牢牢地印在腦子裏。我分明記得有張彩圖上畫着一頭劍龍,劍龍身邊立着幾根超大蘿蔔,幾乎和劍龍一般大;圓滾滾的蘿蔔根表面盛開着許多美麗的花朵。
長大一些之後,我就知道恐龍時代不可能有什麼大蘿蔔。我看到的究竟是什麼東西?但那時不要說那本畫冊,就連圖書室和“單位”都已不復存在,根本無從查證了。這謎團困擾了我差不多20年,甚至讓我懷疑記憶的真實性。直到後來我在另一本書裏看到這張圖:
照片攝自[1]。
我於是釋然,又不禁失笑。記憶確實是真的。只不過這是一類形似蘇鐵的植物。我生在北方,小時候沒見過蘇鐵,就把聚生在樹頂的羽葉當成了蘿蔔纓,膨大的樹幹當成了蘿蔔根,鬧了個“開花大蘿蔔”的笑話。
帶着兒時的溫馨記憶,這一回,井蛙就來介紹一下這類被稱爲本內蘇鐵(Bennettitales)的中生代植物。
Bennettitales以英國植物學家約翰·約瑟夫·本內(John Joseph Bennett,1801–1876)的姓氏命名,中文譯爲“本內蘇鐵目”,顯示它們和延續到現代的蘇鐵目(Cycadales)有親緣關係。這兩類植物曾經合稱爲“蘇鐵植物(Cycadophytes)”,或“莖基植物(Caudophytes)”。不過隨着研究深入,學界逐漸傾向認爲二者屬於不同的演化支,它們的相似性其實是趨同演化的結果。
園藝種植的現代蘇鐵。圖片來源自[2]。

白堊紀,一羣恐龍徜徉在本內蘇鐵森林中。圖片來源自[3]。
可以看到,有些本內蘇鐵的外形確實酷似現代的蘇鐵。它們也長有粗壯的莖幹,在莖幹頂部聚生大叢羽葉。隨着植物生長,老舊葉片枯萎脫落,在莖幹表面留下螺旋排布的菱形葉柄痕。大多數本內蘇鐵葉子都是許多小葉片沿着一根主葉柄兩側對稱或者交錯排布,這樣的複合葉稱爲“一回羽狀複葉(simple pinnate leaf)”。


英國埃克斯伯恩山谷(Ecclesbourne Valley)岩層保留出的本內蘇鐵樹幹化石。鱗片狀結構就是樹幹表面的葉痕。這裏屬於威爾登植物化石羣(Wealden flora),保存了大量早白堊世動植物化石。圖片來源自[4]。

美國北部發現的白堊紀本內蘇鐵——達科塔擬蘇鐵Cycadeoidea dacotensis莖幹化石。此標本高約45釐米,完美保存了胖墩墩的樹幹外形和表皮上的葉痕結構。圖片來源自[5]。
目前發現的本內蘇鐵目植物,高矮胖瘦,分杈不分杈的都有。那些一根矮胖樹墩子上頂着一大蓬葉子,外形最像現代蘇鐵(或者我記憶裏的大蘿蔔)的種類,大體上屬於擬蘇鐵科(Cycadeoideaceae)。
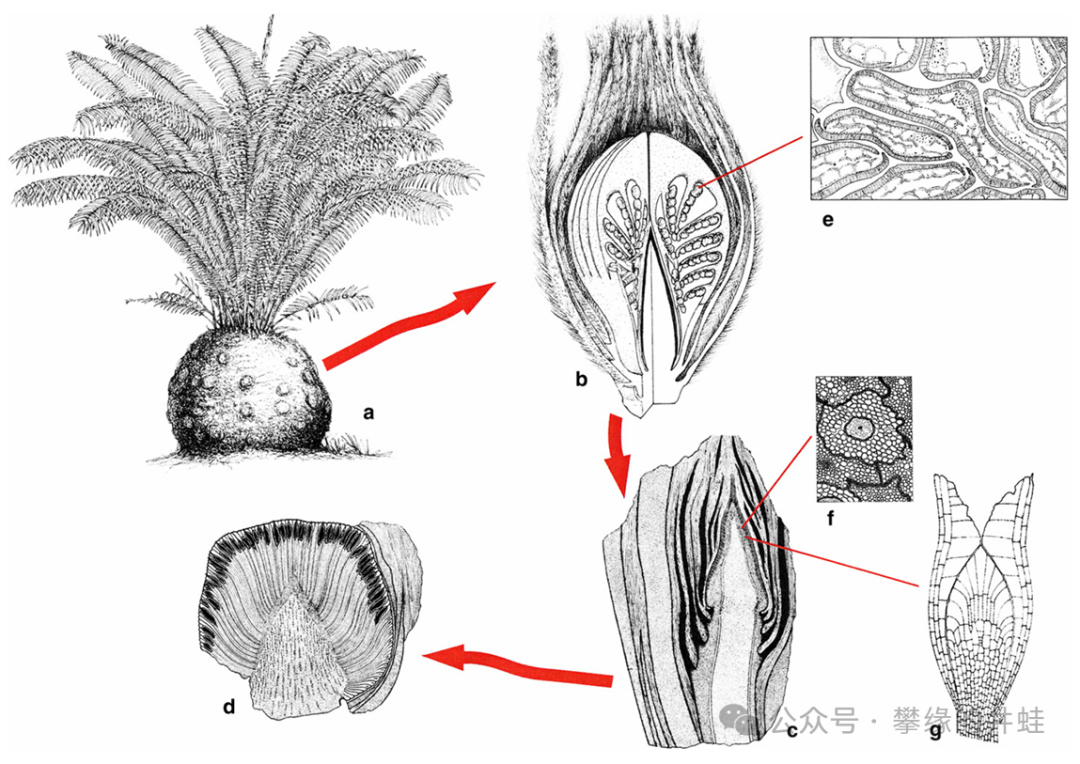
擬蘇鐵科植物的典型特徵。圖片來源自[6]。
另一些樹形比較高挑,樹幹上有少量分枝的種類,被歸入Williamsoniaceae——文獻翻譯並不統一,有叫魏蘭德科、威廉姆遜科、威氏蘇鐵科的。本文暫且稱爲威廉蘇鐵科。
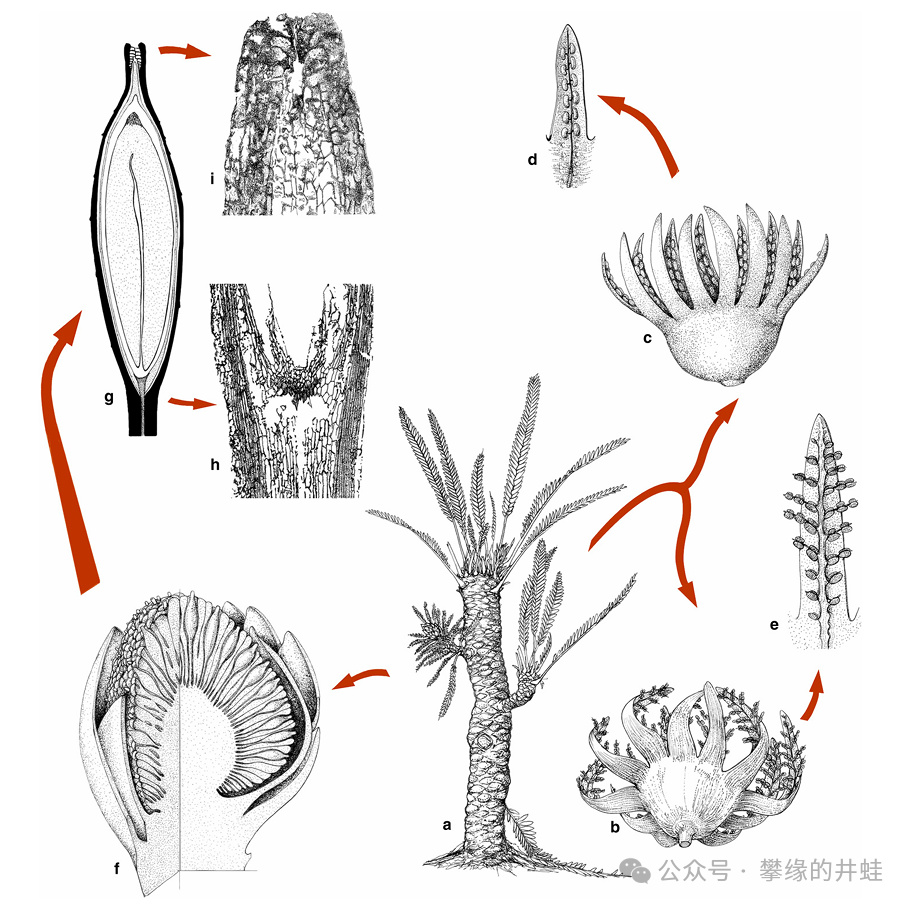
威廉蘇鐵科植物的典型特徵。這個科的外形相對來說不那麼像蘇鐵。圖片來源自[6]。
也有研究者把威廉蘇鐵科當中的一些種類拿出來單獨建立了一個科:Wielandiellaceae,文獻中較少提及。從發音來看,似乎這才應該翻譯成“魏蘭德科”。本文暫且稱爲“維蘭蘇鐵科”。但其實,“維蘭蘇鐵”的外形已經很難和“蘇鐵”聯繫在一起了。

本內蘇鐵三科外形對比。從左到右:1爲維蘭蘇鐵科;2爲威廉蘇鐵科;3爲擬蘇鐵科。圖片來源自[7]。
上圖中除了對比本內蘇鐵的株形,還特別展示了它們的繁殖器官。這也是本內蘇鐵最爲獨特之處。
相信很多朋友都見過鐵樹開花。我們先來看看蘇鐵的繁殖器官。這是最常見的園藝品種,琉球蘇鐵Cycas revoluta:
圖片來源自網絡。
南方綠化帶種植的蘇鐵其實每年都會開花結實。雄性蘇鐵會在樹頂的羽葉叢中央,直立起一根金黃色“大玉米棒”似的雄球花,或者叫小孢子葉球。如果是雌性蘇鐵的話,就在同樣的位置長出一大團像黃色捲心菜似的雌球花,或者叫大孢子葉球。種子就暴露生長在大孢子葉球上(參見第二百三十五回綠之巨人傳9:薪火)。
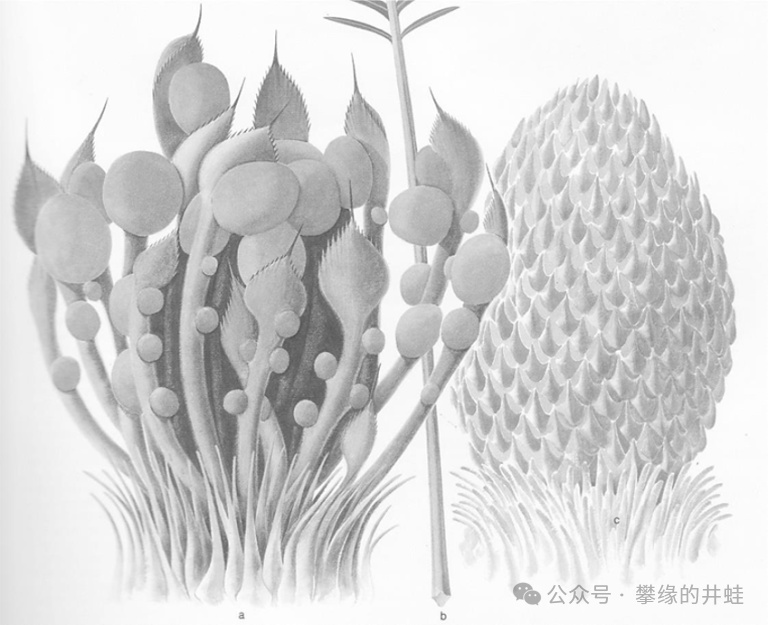
蘇鐵的雌球花(左)和雄球花(右)結構。無論雌雄,組成孢子葉球的每片孢子葉形態都完全相同。雌球花底部的小球就是發育成種子的胚珠(ovule),直接暴露在孢子葉上。圖片來源自網絡。
無論形狀還是結構,這樣的繁殖器官都和我們熟悉的“花”相去甚遠。
本內蘇鐵的情況完全不同。首先,本內蘇鐵的繁殖器官看上去非常像是真正的“花”。葉球內外層的孢子葉(往往也是雄性的那部分)形態分化,外層特化成放射狀的苞片(bracts),形成類似被子植物花瓣的結構。
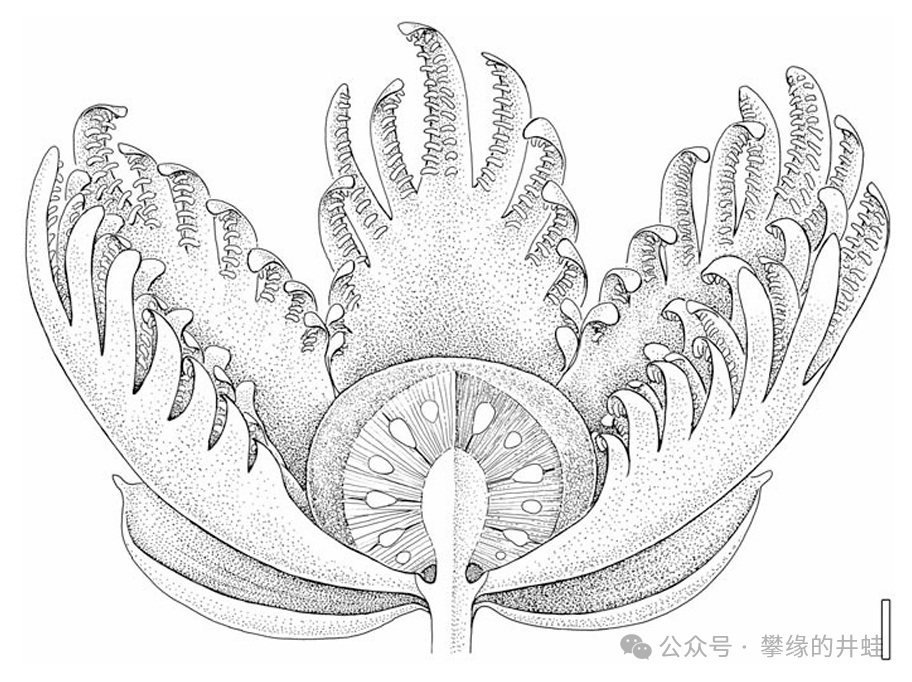
根據幾種三疊紀本內蘇鐵化石復原的早期本內蘇鐵“花朵”結構。外層花瓣狀的是產生花粉的小孢子葉球。中心的球狀結構是接受花粉的大孢子葉球,內部含有發育成種子的胚珠。圖片來源自[8],標尺長度10毫米。
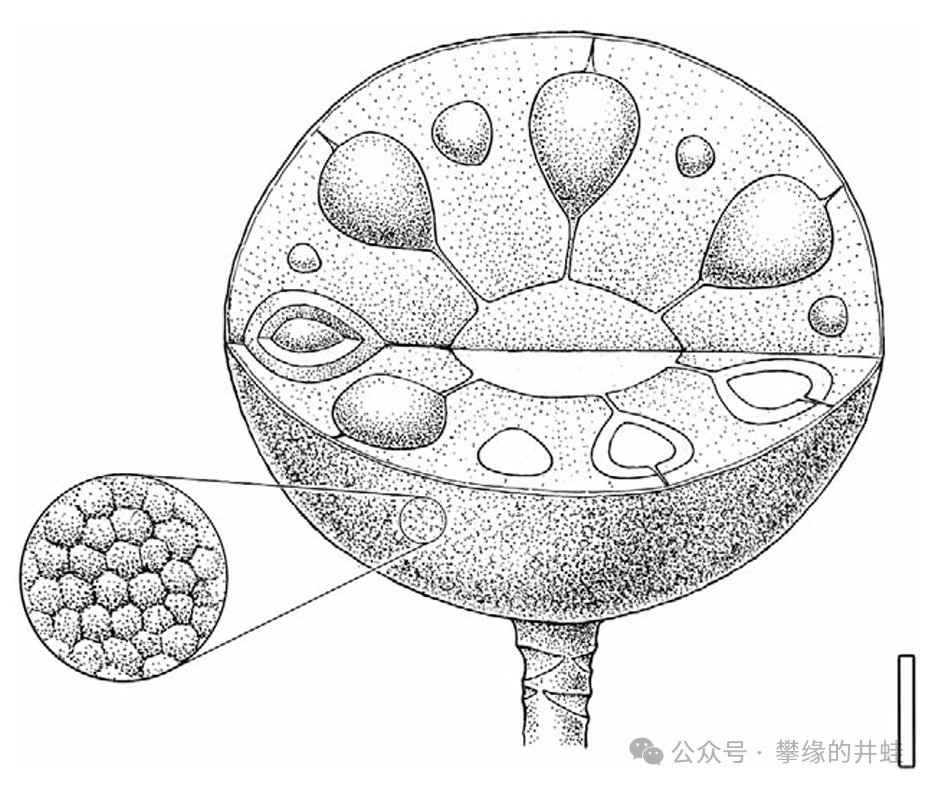
胚珠受精後,大孢子葉球就會逐漸發育成類似松塔的球果,或者叫“種實(seed cone)”。圖片來源自[8],標尺長度10毫米。
像本內蘇鐵這種,同一朵“花”裏既有產生花粉的雄性結構(小孢子葉),也有孕育種子的雌性結構(大孢子葉),稱爲“兩性花(bisexual flower)”。蘇鐵只有“單性花(unisexual flower)”,這也是二者的主要區別。
在烏拉爾山的羣芳凋零之後(參見第二百十一回似花還似非花),地球上又一次綻放出美麗的花朵。

澳大利亞發現的三疊紀本內蘇鐵花朵化石。a:Williamsonia eskensis sp. nov.;b:Williamsonia ipsvicensis sp. nov.。圖片來源自[9],標尺長度10毫米。

伊朗發現的晚三疊世本內蘇鐵花朵Williamsonia sp.。圖片來源自[5]。
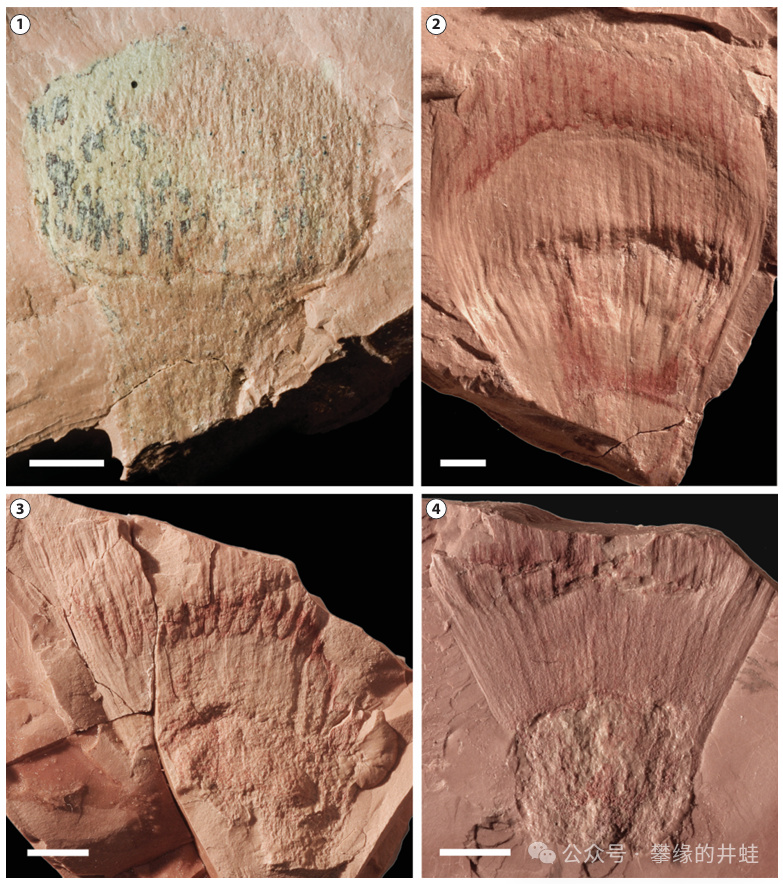
巴西南部發現的早侏羅世本內蘇鐵Williamsonia potyporanae sp. nov.花朵化石。圖片來源自[10],標尺長度5毫米。

Williamsonia potyporanae sp. nov.花朵結構復原圖。1、2展示它外層的兩層苞片,3、4展示胚珠生長和種子發育。圖片來源自[10]。

英國發現的中侏羅世本內蘇鐵花朵Weltrichia spectabilis。圖片來源自[5]。

日本東北部發現的晚侏羅世本內蘇鐵Kimuriella densifolia。着重展示它的花朵形態。圖片來源自[11],標尺長度1釐米。

同時保存了枝條、羽葉、花朵(A圖黑色箭頭、C圖)和球果(A圖白色箭頭、B圖)的Kimuriella densifolia化石。圖片來源自[11],標尺長度1釐米。
蘇鐵的球花只生長在莖幹頂端,被羽葉簇擁環繞。本內蘇鐵的開花位置則更加多樣。有的種類像蘇鐵一樣長在枝幹頂端。

瑞士發現的一種晚三疊世維蘭蘇鐵Wielandiella angustifolia化石。包括a枝幹,b花朵,c球果,d葉片。圖片來源自[12],標尺長度:a圖10釐米,其它1釐米。

Wielandiella angustifolia復原圖。它的花朵就長在每根分枝的頂端,圍繞一圈葉片。圖片來源自[12]。
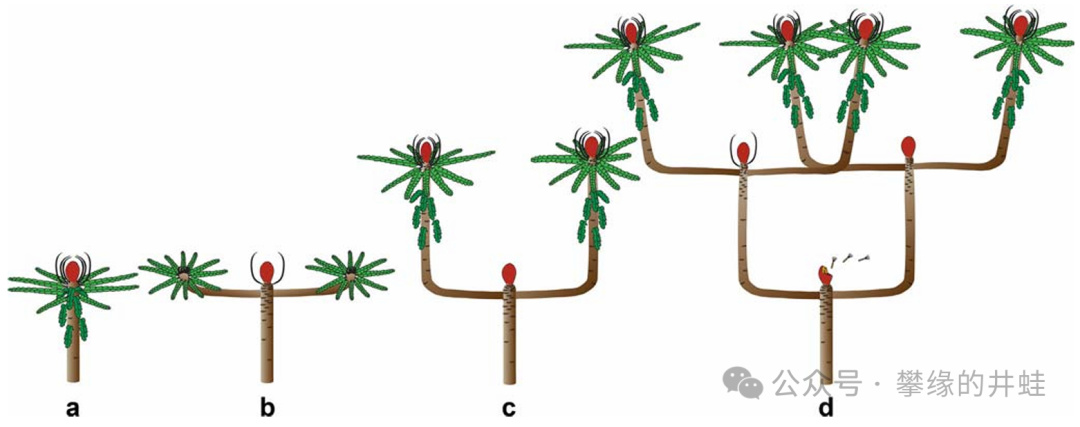
本圖展示了一些威廉蘇鐵和維蘭蘇鐵奇特的生長模式。每根枝條頂端開花結實,然後分生出兩根次級枝條。不斷重複“頂端開花”-“花周生葉”-“兩歧分生”的過程。圖片來源自[12]。
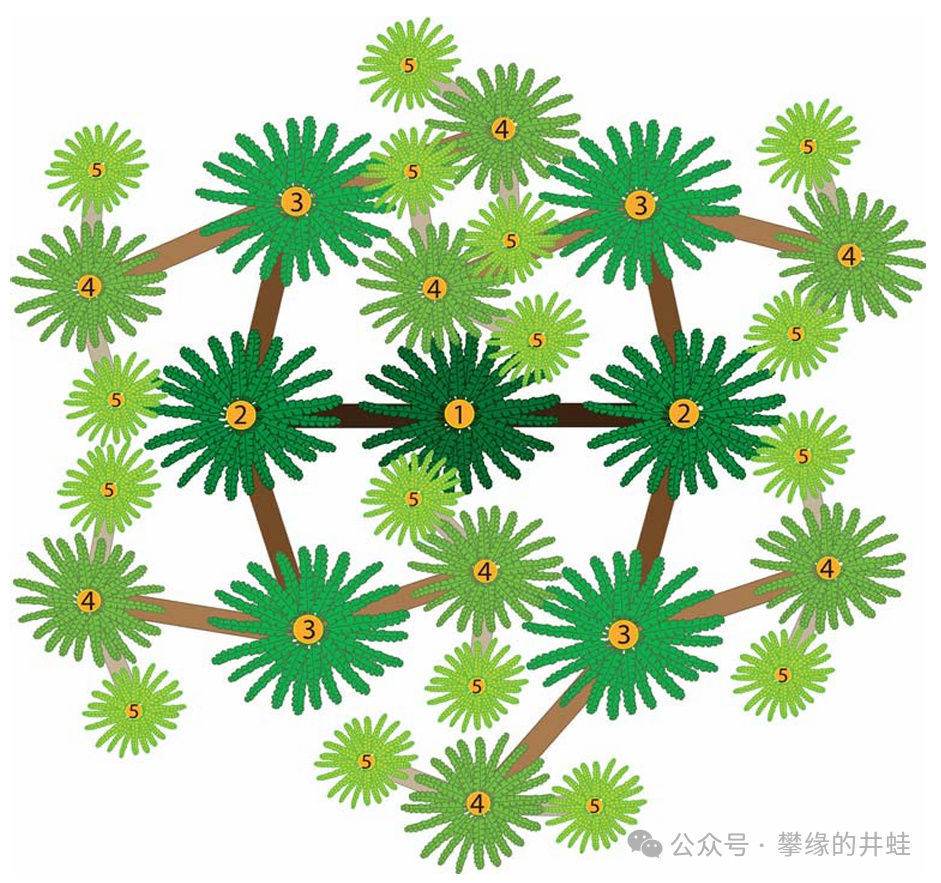
這種分形生長可以帶來一個好處。從株頂俯視(如上圖),各級枝條頂端的葉叢形成類似鑲嵌的構圖,相互錯開,避免遮擋,可以最大限度地接受陽光。演化的力量真是神奇。圖片來源自[12]。
有在枝頂開花的,也有在葉柄基部與莖的連接處,或者叫“葉腋”處開花的。
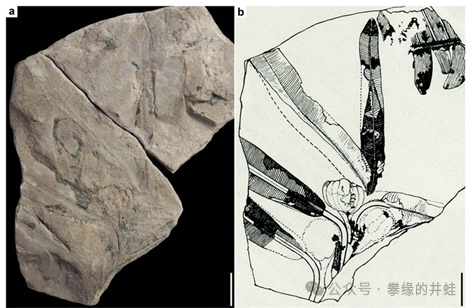
英國發現的侏羅紀本內蘇鐵:Williamsoniella coronata化石。圖片來源自[12],標尺長度1釐米。

Williamsoniella coronata復原圖。相比“蘇鐵”,它的形態更像一些草本植物,比如花生在葉腋的部位;又比如組成“羽葉”的小葉片癒合,形成類似整片條形葉的結構。圖片來源自[12]。
最像蘇鐵的擬蘇鐵科,也把花開在葉腋的位置,只不過開花時葉片已經脫落了。一眼看上去,就像矮墩墩的樹幹上開滿了花,像個臭美的大胖腦袋。
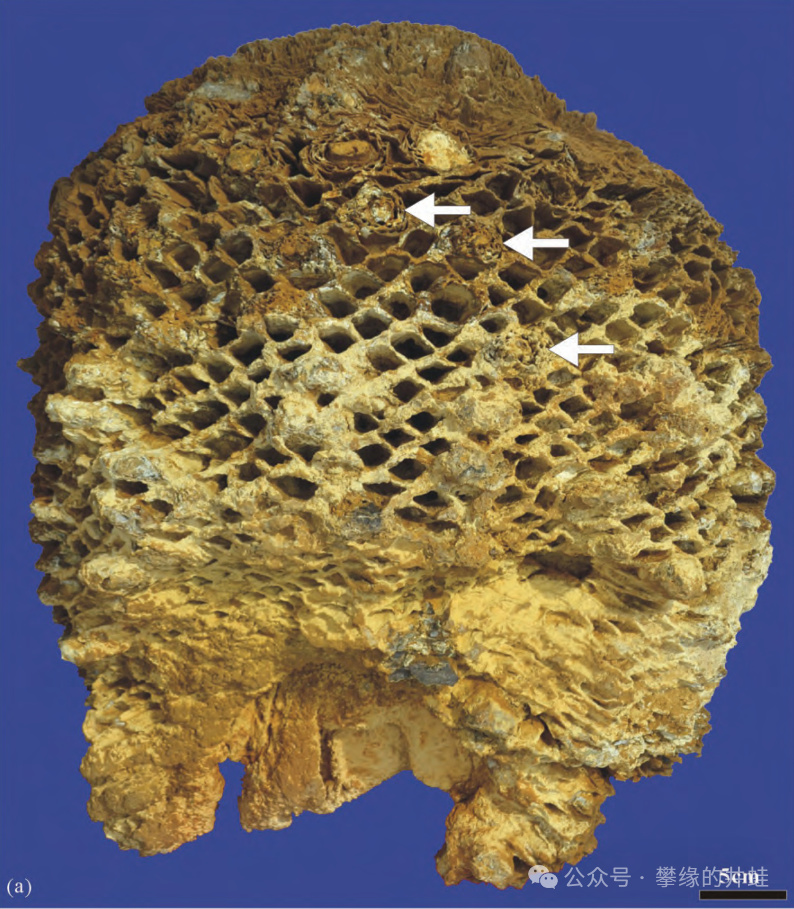
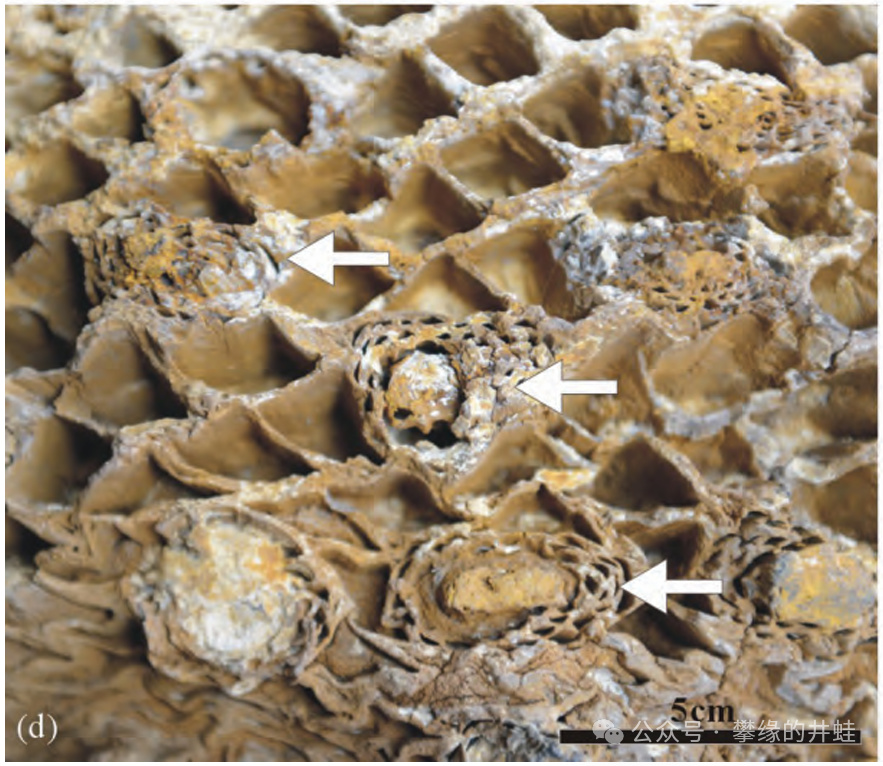
內蒙古包頭達爾罕茂明安聯合旗發現的白堊紀擬蘇鐵屬Cycadeoidea莖幹化石。箭頭指示散生於葉柄基之間的球果。每個球果都曾經是一朵花。圖片來源自[13]。
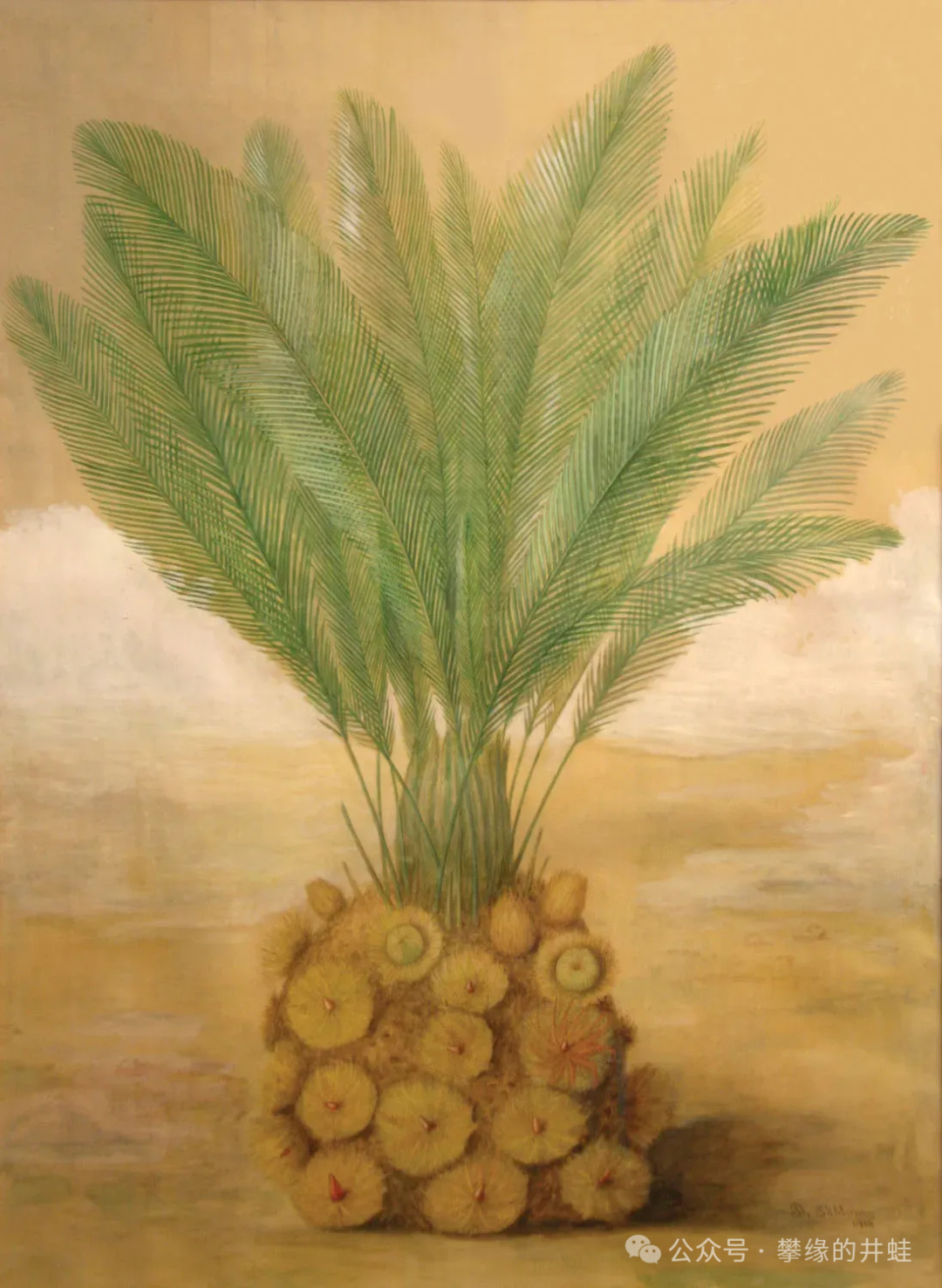
擬蘇鐵復原圖。這就是我記憶中的“開花大蘿蔔”真正的樣子。現在再看,又有點像蘇鐵和仙人掌的融合體。圖片來源自網絡。
本內蘇鐵的“花”和被子植物的太像了,曾經有假說認爲被子植物起源於本內蘇鐵。不過現在已經被否定了。雖然它們的“花”相似,但本內蘇鐵畢竟屬於“裸子植物”,只有種子,不結果實,完全不同於被子植物。
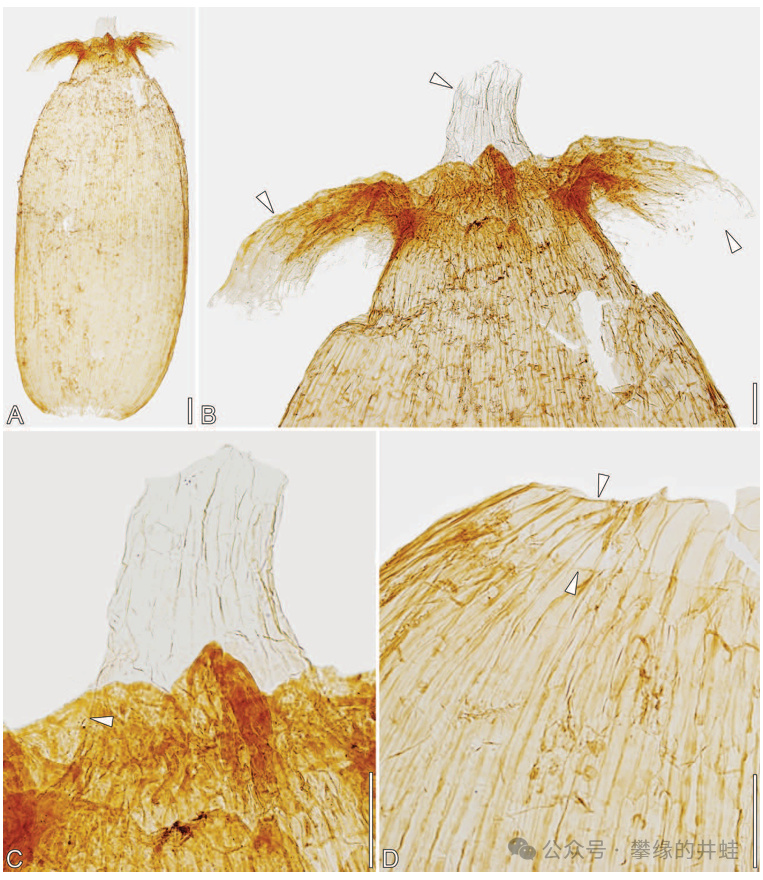
奧地利發現的晚三疊世本內蘇鐵Westersheimia pramelreuthensis。上圖展示了它的胚珠結構,沒有子房壁包裹,是裸露的。這也是所有“裸子植物”的共同特徵。圖片來源自[14],標尺長度 A-C爲100微米,D爲10微米。
Westersheimia pramelreuthensis的種子結構。由於沒有子房壁發育成的果皮,它也就只有種子,沒有果實。圖片來源自[14] ,標尺長度200微米。
雖然本內蘇鐵不再被當做被子植物的祖先,但研究者仍然認爲它們和被子植物的親緣關係遠比其他裸子植物更近,甚至認爲本內蘇鐵可以和被子植物一起歸入顯花植物(Anthophytes)。

研究者設想的原始花朵形態,具有這種花朵的古老植物的後代包括被子植物、本內蘇鐵和買麻藤目(Gnetales)等。圖片來源自[15] 。
相比樹幹、花朵和種子,本內蘇鐵留下了更豐富的葉片化石。本內蘇鐵的羽狀複葉形態多樣,經常直接拿來給化石種命名,比如側羽葉屬Pterophyllum、異羽葉屬Anomozamites、耳羽葉屬Otozamites、網羽葉屬Anthrophyopsis、毛羽葉屬Ptilophyllum等等。

德國南部雷塔恩植物化石羣(Rhaetian flora)發現的三疊紀末到侏羅紀早期側羽葉屬化石。圖片來源自[16],標尺長度1釐米。

德國南部雷塔恩植物化石羣(Rhaetian flora)發現的三疊紀末到侏羅紀早期本內蘇鐵葉片化石。1-7爲側羽葉屬,8、9爲異羽葉屬。圖片來源自[16],標尺長度1釐米。

澳大利亞發現的晚白堊世本內蘇鐵葉片化石。5a爲毛羽葉屬,5b、5c爲耳羽葉屬。圖片來源自[17]。
一些中國發現的異羽葉屬種葉片形態。中生代本內蘇鐵之繁盛,可見一斑。圖片來源自[18]。
本內蘇鐵目在早二疊世便已出現,此後緩慢發展,並在二疊紀末大滅絕中倖存。卡尼期洪積事件徹底改變了陸地植物版圖。適應溼熱環境的本內蘇鐵迅速崛起,自晚三疊世到中白堊世,它們都是構築陸地生態系統的堅固基石。
約旦發現的晚二疊世本內蘇鐵Pterophyllum pottii化石。圖片來源自[19],標尺長度:A,B爲1釐米,C爲5毫米。
A-G:約旦發現的晚二疊世本內蘇鐵Nilssoniopteris jogiana。H-Q:中國發現的早二疊世本內蘇鐵山西肋羽葉Nilssoniopteris shanxiensis。圖片來源自[19],標尺長度:A = 1 cm; B = 5 mm; C,D = 100 μm; E = 200 μm; F-H = 20 μm; J = 50 μm; K = 100 μm; L = 50 μm; M = 20 μm; N,O = 200 μm; P-R = 20 μm。
從上面列舉的化石不難看出,本內蘇鐵目在中生代多樣性極高,從高大喬木到低矮灌木,到柔弱草本,佔據了極其廣闊的生態位。自然也養活了不計其數的動物,尤其是各種植食昆蟲。

內蒙古寧城道虎溝侏羅紀狹長焦帶羽葉Nilssoniopteris longifolius上的昆蟲取食痕跡。a、b箭頭指示啃咬痕跡,c箭頭指示刺吸痕跡。圖片來源自[20] ,標尺長度:a爲2毫米,b爲1毫米,c爲2毫米。
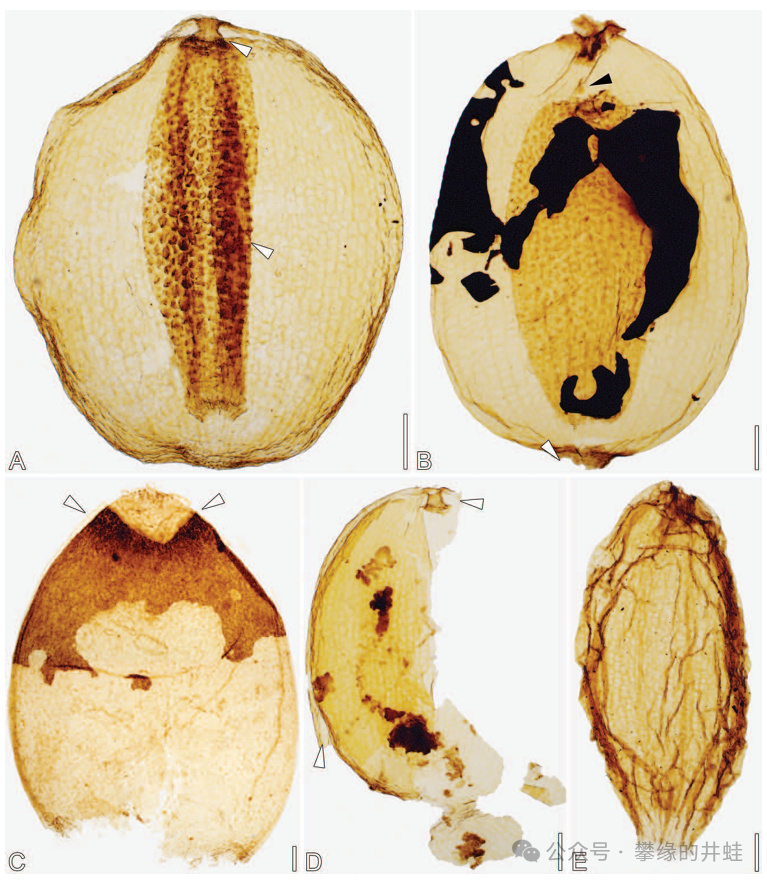
奧地利發現的晚三疊世Nilssoniopteris haidingeri葉片化石,上面發現了昆蟲的卵殼(長度約200微米)。圖片來源自[5]。

內蒙寧城道虎溝發現的中侏羅世具毛異羽葉Anomozamites villosus化石。這種本內蘇鐵簡直被當時的昆蟲們當成了集體食堂,研究者在它的身上發現了多達37種損傷,包括但不限於:邊緣取食、孔洞取食、表面取食、留脈式取食、刺吸取食、產卵、潛葉、造癭、種子取食、鑽蛀,以及真菌病害侵染等等。圖片來源自[12],標尺長度:a, c, d = 1cm, b = 5mm, e, f = 2 mm。
化石證據記錄了異羽葉Anomozamites植物上多樣的植食方式。圖片來源自[21]。
具毛異羽葉上的18類昆蟲植食復原圖。圖片來源自[21]。
有理由相信,本內蘇鐵華麗的“花朵”,也應該是和昆蟲互動的結果。很可能有一些昆蟲在取食葉片或花球的過程中,和本內蘇鐵協同演化,從食客逐漸演變成傳播花粉的使者。本內蘇鐵特化的苞片,就是爲了招徠它們專門演化的看板。可惜目前還沒有發現這些傳粉昆蟲的化石證據,我們不妨耐心等待。
有的昆蟲喫着喫着,把自己也變成了本內蘇鐵的樣子。還是在內蒙古寧城道虎溝生物羣,研究者發現有三種直翅目鳴螽的前翅演化出極其類似異羽葉的色斑。
侏羅紀擬態型鳴螽的前翅化石。注意上面的色塊形狀。圖片來源自[22]。
這是鳴螽們努力模仿的異羽葉化石。圖片來源自[22]。
棲息在異羽葉上的擬態鳴螽復原圖。這些小蟲也是各顯其能。有一片翅膀上畫一張羽葉的;也有每片翅膀畫一半,合攏後在背上拼出一張羽葉的。圖片來源自[22]。
這些昆蟲拼命讓自己融入背景,自然是爲了躲避天敵的捕食。那麼它們的天敵是誰呢?在侏羅紀中期,鳥類、蝙蝠、樹蛙、避役等等現代食蟲動物還都沒有出現。鳴螽們的主要敵人,是小型翼龍、樹棲恐龍,以及各種哺乳型的鼠輩。
內蒙古寧城五化地區發現的侏羅紀哺乳形類:微小柱齒獸Microdocodon gracilis。它正在一株本內蘇鐵(看樣子是某種維蘭蘇鐵)的枝杈間捕食小昆蟲。圖片來源自[23]。
在合弓獸們最落魄不堪的歲月裏,本內蘇鐵溫柔地爲這些毛絨絨鬧哄哄的小東西提供了棲身之所和果腹食物。
不知讀到這裏的朋友們是否聯想到了別的什麼植物。我一路寫下來,腦海裏一直在把本內蘇鐵和銀杏(Ginkgo)做對比。它們全都起源古老,都在中生代崛起成爲最繁盛的陸生植物類羣,都供養起一套基於自己的複雜食物網;它們都在竭力突破“裸子植物”的底層限制,優化自己的葉片、花朵和種子;它們都是恐龍的背景板,也都曾呵護過柔弱的哺乳動物;甚至人們發現的極少數中生代擬態昆蟲,也分別擬態了本內蘇鐵和銀杏的葉子。何其相似,何其相似——包括它們之後的命運。
白堊紀中期,被子植物席捲全球。這些真正的花花果果很快就把本內蘇鐵(當然還有銀杏)打得一潰千里。以至於自晚白堊世後大約6000萬年的時間裏,人們在地層中都找不到本內蘇鐵的葉片、莖幹、花朵、種子等“宏體化石”,只有一些零星的花粉記錄,證明它們仍然頑強地生存着。
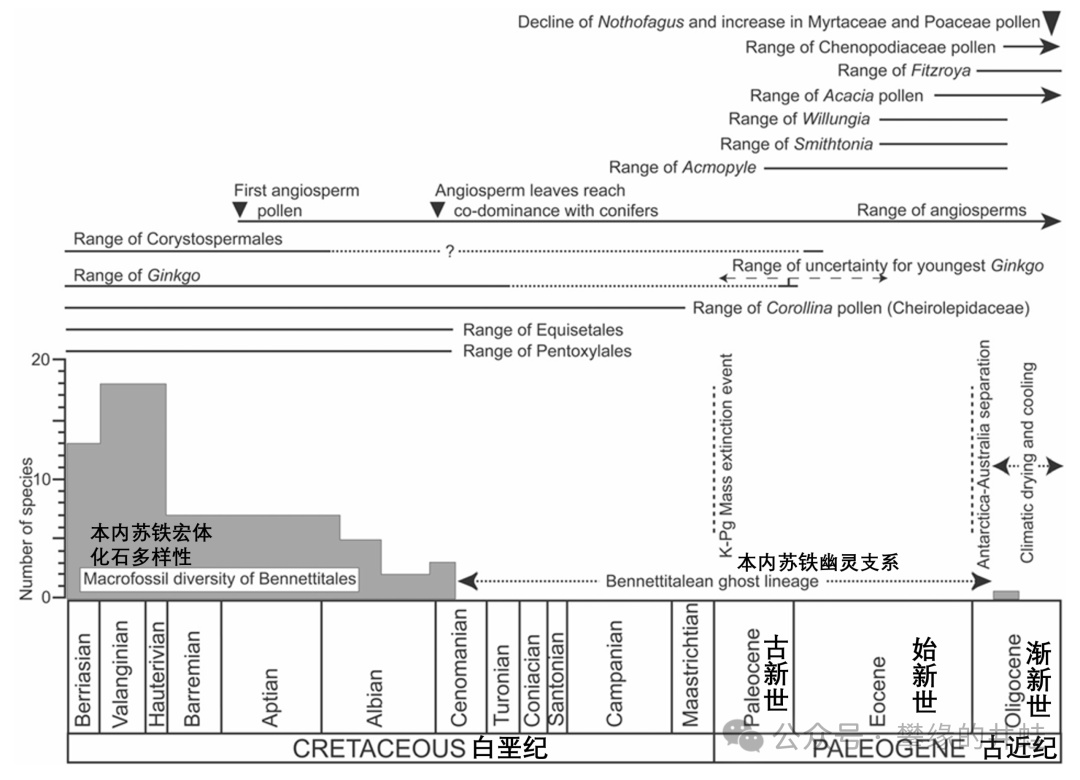
本內蘇鐵和其他相關植物(包括銀杏)的化石統計。在白堊紀森諾曼期後,本內蘇鐵的化石就再難尋覓。圖片來源自[24]。
這些殘存的本內蘇鐵挺過了白堊紀末大滅絕,它們跌跌撞撞地走進了新生代,見證了自己曾經護佑的小哺乳動物豹變成爲這顆星球新的主宰。然而隨着冰期來臨,全球氣溫驟降,適應溫熱環境的本內蘇鐵越來越難容身於這個冷酷的新世界。目前人們發現的最後的本內蘇鐵化石,停留在漸新世。

澳大利亞新南威爾士(A-H)和塔斯馬尼亞(I-L)發現的漸新世本內蘇鐵:Ptilophyllum muelleri葉片化石。它們是這個古老族裔的最後身影。圖片來源自[24],標尺長度:A-J = 10mm,K, L = 5 mm。
自此之後,本內蘇鐵的身影,從地球上徹底消失了。本內蘇鐵終究還是沒能有銀杏那般的幸運。這對苦命鴛鴦還是沒能攜手走到最後。神奇的“開花大蘿蔔”,還有它們對生命形態的探索,都在3000萬年前畫上了終止符。
地球名片
生物分類:植物界-維管植物-蘇鐵植物門-蘇鐵綱-本內蘇鐵目(或獨立爲本內蘇鐵綱)
存在時間:早二疊世?至漸新世(繁盛於晚三疊世至白堊紀中期,白堊紀末基本滅絕,極少數種類殘存到漸新世)
現存種類:無
化石種類:未找到完整統計
生活環境:陸地(喜溫溼環境)
代表特徵:形態多樣。葉一般爲一回羽狀複葉,具雙脣型氣孔;孢子葉球結構複雜,類似被子植物的花,且大多數爲兩性花
代表屬種:擬蘇鐵屬、威廉蘇鐵屬、維蘭蘇鐵屬、側羽葉屬、異羽葉屬、耳羽葉屬、網羽葉屬、毛羽葉屬等
參考文獻
[1] 劉後一,陳淳,王幼於,生物是怎樣進化的,中國青年出版社,1984
[2] https://www.gardeningknowhow.com/ornamental/foliage/cycads/growing-cycad-plants.htm
[3] https://www.newgeology.us/presentation14.html
[4] Peter Austen, Bennettitale trunks from Hasting. Wealden News No.8, February 2010
[5] Stephen McLoughlin, Christian Pott, Harvesting the extinct Bennettitales.
[6] Gar W. Rothwell, William L. Crepet, Ruth A. Stockey, IS THE ANTHOPHYTE HYPOTHESIS ALIVE AND WELL? NEW EVIDENCE FROM THE REPRODUCTIVE STRUCTURES OF BENNETTITALES. American Journal of Botany 96(1): 296–322. 2009, doi:10.3732/ajb.0800209
[7] http://www.evolution-biologique.org/index.php/conquete-des-continents/histoire-des-v%c3%a9g%c3%a9taux-terrestres/groupes-v%c3%a9g%c3%a9taux-terrestres/bennettitales.html?page=histoire-de-la-vie/conquete-des-continents/histoire-des-v%c3%a9g%c3%a9taux-terrestres/groupes-v%c3%a9g%c3%a9taux-terrestres/bennettitales.html
[8] Christian Pott, Michael Krings, Hans Kerp, Else Marie Friis, Reconstruction of a bennettitalean flower from the Carnian (Upper Triassic) of Lunz, Lower Austria. Review of Palaeobotany and Palynology 159 (2010) 94–111, doi: 10.1016/j.revpalbo.2009.11.004
[9] Stephen McLoughlin, Christian Pott, Ian H. Sobbe, The diversity of Australian Mesozoic bennettitopsid reproductive organs. Palaeobio Palaeoenv (2018) 98: 71–95, DOI: 10.1007/s12549-017-0286-z
[10] RONALDO BARBONI, TÂNIA LINDNER DUTRA, NEW “FLOWER” AND LEAVES OF BENNETTITALES FROM SOUTHERN BRAZIL AND THEIR IMPLICATION IN THE AGE OF THE LOWER MESOZOIC DEPOSITS. AMEGHINIANA - 2013 - Tomo 50 (1): 14–32
[11] Pott, Christian, and Takimoto, Hideo, Kimuriella gen. nov. (Bennettitales), a Whole-Plant Bennettite from the Oxfordian (Upper Jurassic) Tochikubo Formation of Shidazawa, Minamisōma, Fukushima Prefecture, Northeast Japan. Paleontological Research, 26(2) : 158-186, https://doi.org/10.2517/PR200020
[12] Christian Pott, Stephen McLoughlin, Divaricate growth habit in Williamsoniaceae (Bennettitales): unravelling the ecology of a key Mesozoic plant group. Palaeobio Palaeoenv, DOI: 10.1007/s12549-014-0157-9
[13] Cheng Y M, Wang X, Yang X N, Tan K, Jin Y G. The first discovery of a Cretaceous big Cycadeoidaceae (Bennettitales) trunk from Inner Mongolia. Geological Bulletin of China, 2016, 35(8):1230-1233
[14] Christian Pott, WESTERSHEIMIA PRAMELREUTHENSIS FROM THE CARNIAN (UPPER TRIASSIC) OF LUNZ, AUSTRIA: MORE EVIDENCE FOR A UNITEGMIC SEED COAT IN EARLY BENNETTITALES. Int. J. Plant Sci. 177(9):771–791. 2016
[15] Gar W. Rothwell, Ruth A. Stockey, Independent evolutIon of Seed enclosure In the BennettItales: evIdence from the anatomIcally preserved cone Foxeoidea connatum gen. et sp. nov.
[16] Pott, Christian, Schmeißner, Stefan, Dütsch, Günter, Van Konijnenburg-van Cittert, Johanna H.A., Bennettitales in the Rhaetian flora of Wüstenwelsberg, Bavaria, Germany, Review of Palaeobotany and Palynology (2016), doi: 10.1016/j.revpalbo.2016.04.01
[17] Steve McLoughlin, Christian Pott, On the Trail of Australia’s Youngest Bennettitales.
[18] Xu, X., Gou, X.D., Wan, S., Liu, H.Y., Wei, H.B., Liu, J.R., Ding, J.H., Yang, B., Shu, Y.P., Hua, Y.P., Feng, Z.*, 2019. Anomozamites (Bennettitales) in China: species diversity and temporal-spatial distribution. Palaeontographica Abteilung B,300(1–6): 21–46. DOI:10.1127/palb/2019/0067
[19] Blomenkemper P, Bäumer R, Backer M, Abu Hamad A, Wang J, Kerp H and Bomfleur B (2021) Bennettitalean Leaves From the Permian of Equatorial Pangea—The Early Radiation of an Iconic Mesozoic Gymnosperm Group. Front. Earth Sci. 9:652699. doi: 10.3389/feart.2021.652699
[20] Xin Wang, Qiaoling Ding, Artai A. Santos, Torsten Wapplere, Nilssoniopteris longifolius Chang from the Middle–Late Jurassic of China: Implications for Bennettitales-insect interactions. Review of Palaeobotany and Palynology 297 (2022) 104582, https://doi.org/10.1016/j.revpalbo.2021.104582
[21] http://www.ivpp.cas.cn/klt/kycg_home/202412/t20241212_7454563.html
[22] https://nigpas.cas.cn/kyjz/202509/t20250902_7957827.html
[23] https://www.sci.news/paleontology/microdocodon-gracilis-07423.html
[24] Stephen McLoughlin, Raymond J. Carpenter, Christian Pott, PTILOPHYLLUM MUELLERI (ETTINGSH.) COMB. NOV. FROM THE OLIGOCENE OF AUSTRALIA: LAST OF THE BENNETTITALES? Int. J. Plant Sci. 172(4): 574–585. 2011., DOI: 10.1086/658920
更多遊戲資訊請關註:電玩幫遊戲資訊專區
電玩幫圖文攻略 www.vgover.com












![[6.27]夏促來襲!超多骨折!百款史低新史低這次你一定要入庫!](https://imgheybox1.max-c.com/web/bbs/2026/06/26/bfb8adfbb675422149a024f4b7064470.png?imageMogr2/auto-orient/ignore-error/1/format/jpg/thumbnail/398x679%3E)



